| |
Chapter Thirteen | Delgado Index | Chapter Fifteen
Jose Delgado's "Physical Control of the Mind"
Hell and Heaven Within the Brain:
The Systems for Punishment and Reward
When man evolved above other powerful animals, the size and complexity of his brain increased, giving him superior intelligence along with more anguish, deeper sorrow, and greater sensitivity than any other living creature. Man also learned to enjoy beauty, to dream and to create, to love and to hate. In the education of children as well as in the training of animals, Punishment and reward constitute the most powerful motivations for learning. In our hedonistic orientation of life to minimize pain and seek pleasure, we often attribute these qualities to the environment without realizing that sensations depend on a chain of events which culminates in the activation of determined intracerebral mechanisms. Physical damage, the loss of a beloved child, or apocalyptic disaster cannot make us suffer if some of our cerebral structures have been blocked by anesthesia. Pleasure is not in the skin being caressed or in a full stomach, but somewhere inside the cranial vault.
At the same time pain and pleasure have important psychic and cultural components related to individual history. Men inhibited by sortie extraordinary tribal or religious training to endure discomfort have been tortured to death without showing signs of suffering. It is also known that in the absence of physical injury, mental elaboration of information may produce the worst kind of suffering. Social rejection, guilt feelings, and
other personal tragedies may produce greater autonomic, somatic, and psychological manifestations than actual physical pain.
There is strong reluctance to accept that such personal and refined interpretations of reality as being afraid and being in love are contingent on the membrane depolarization of determined clusters of neurons, but this is one aspect of emotional phenomena which should not be ignored. After frontal lobotomy, cancer patients have reported that the pain persisted undiminished, but that their subjective suffering was radically reduced, and they did not complain or request as much medication as before surgery. Lobotomized patients reacted to noxious stimuli as much, if not more, than before their operations, jumping at pinpricks and responding quickly to objective tests of excessive heat, but they showed decreased concern. It seems that in the frontal lobes there is a potentiating mechanism for the evaluation of personal suffering, and after lobotomy the initial sensation of pain is unmodified, while the reactive component to that feeling is greatly diminished. This mechanism is rather specific of the frontal lobes; bilateral destruction of the temporal lobes fails to modify personal suffering.
Important questions to resolve are: Do some cerebral structures have the specific role of analyzing determined types of sensations? Is the coding of information at the receptor level essential for the activation of these structures Not too long ago, many scientists would have dismissed as naive the already demonstrated fact that punishment and reward can be induced at will by manipulating the controls of an electrical instrument connected to the brain.
Perception of Suffering
In textbooks and scientific papers, terms such as "pain receptors," "pain fibers," and "pain pathways" are frequently used, but it should be clarified that peripheral nerves do not carry sensations. Neuronal pathways transmit only patterns of elec-
trical activity with a message that must be deciphered by the central nervous system, and in the absence of brain there is no pain, even if some reflex motor reactions may still be present. A decapitated frog cannot feel but will jump away with fairly good motor coordination when pinched in the hind legs. During competitive sports or on the battlefield, emotion and stress may temporarily block the feeling of pain in man, and often injuries are not immediately noticed. The cerebral interpretation of sensory signals is so decisive that the same stimulus may be considered pleasant or unpleasant depending on circumstances. A strong electrical shock on the feet scares a dog and inhibits its secretion of saliva. If, however, the same "painful" excitation is followed for several days by administration of food, the animal accepts the shock, wagging its tail happily and salivating in anticipation of the food reward. Some of these dogs have been trained to press a lever to trigger the electric shock which preceded food. During sexual relations in man, bites, scratches, and other potentially painful sensations are often interpreted as enjoyable, and some sexual deviates seek physical punishment as a source of pleasure.
The paradox is that while skin and viscera have plentiful nerve endings for sensory reception, the brain does not possess this type of innervation. In patients under local anesthesia, the cerebral tissue may be cut, burned, pulled apart, or frozen without causing any discomfort. This organ so insensitive to its own destruction is, however, the exquisite sensor of information received from the periphery. In higher animal species there is sensory differentiation involving specialized peripheral receptors which code external information into electrical impulses and internal analyzers which decode the circulating inputs in order to give rise to the perception of sensations.
Most sensory messages travel through peripheral nerves, dorsal roots, spinal cord, and medulla to the thalamic nuclei in the brain, but from there we lose their trail and do not know where the information is interpreted as painful or pleasurable, or how affective components are attributed to a sensation (212, 220).
Although anatomical investigations indicate that thalamic fibers project to the parietal "sensory" cortex, stimulation of this area does not produce pain in animals or man. No discomfort has been reported following electrical excitation of the surface or depth of the motor areas, frontal lobes, occipital lobes, cingulate gyros, and rnany other structures, while pain, rage, and fear have been evoked by excitation of the central gray tegmentum, and a few other regions.
Animals share with man the expressive aspect of emotional manifestations. When a dog wags its tail, we suppose it is happy, and when a cat hisses and spits we assume that it is enraged, but these interpretations are anthropomorphic and in reality we do not know the feelings of any animal. Several authors have tried to correlate objective manifestations with sensations; for example, stimulation of the cornea of the eye provokes struggling, pupillary dilatation, and rise of blood pressure (87), but these responses are not necessarily related to awareness of feelings, as is clearly demonstrated by the defensive ability of the decapitated frog. Experimental investigation of the mechanisms of pain and pleasure is handicapped in animals by their lack of verbal communication, but fortunately we can investigate whether an animal likes or dislikes the perceived sensations by analyzing its instrumental responses. Rats, monkeys, and other species can learn to press a lever in order to receive a reward such as a food pellet or to avoid something unpleasant such as an electric shock to the skin. By the voluntary act of instrumental manipulation, an animal expresses whether or not the food, shock, or brain stimulation is desirable, allowing for the objective qualification of the sensation. In this way, many cerebral strictures have been explored to identify their positive or negative reinforcing properties.
At present it is generally accepted that specific areas of the brain participate in the integration of pain sensations, but the mechanism is far from clear, and in our animal experiments we do not know if we are stimulating pathways or higher centers of integration. The concept of a straight conduction of pain
messages from the periphery up to the central nervous system was too elemental. Incoming messages are probably processed at many levels with feedbacks which modify the sensitivity and the filtering of information at many stages including the peripheral receptor level. Brain excitation, therefore, may affect transmission as well is the elaboration of inputs and feedback modulation. Electrical stimuli do not carry any specific message because they are a monotonous repetition of similar pulses, and the fact that they constitute a suitable trigger for central perception of pain means that the reception of a patterned code is not required, but only the nonspecific activation of neuronal pools which are accessible to investigation. In addition to the importance of these studies for finding better therapies for the alleviation of pain, there is another aspect which has great social interest: the possible relations between pain perception and violence.
Violence Within the Brain
The chronicle of human civilization is the story of a cooperative venture consistently marred by self-destruction, and every advance has been accompanied by increased efficiency of violent behavior. Early man needed considerable physical strength -,end skill to defend himself or attack other men or beasts with stones, arrows, or swords, but the invention of explosives and subsequent development of firearms have made unskilled individuals more powerful than mythical warriors of the past. The technology for destruction has now placed at the disposal of man a vast i arsenal of ingenious weapons which facilitate all forms of violence including crimes against property, assassinations, riots, and wars, threatening not only individual life and national I stability but the very existence of civilization.
Ours is a tragically unbalanced industrial society which devotes most of its resources to the acquisition of destructive power and invests insignificant effort in the search which could provide the true weapons of self-defense: knowledge of the
mechanisms responsible for violent behavior. They are necessarily related with intracerebral processes of neuronal activity, even if the triggering causality may reside in environmental circumstances. Violence is a product of cultural environment and is an extreme form of aggression, distinct from modes of self-expression required for survival and development under normal conditions. Man may react to unpleasant or painful stimuli with violence-he may retaliate even more vigorously than he is attacked-but only if he has been taught by his culture to react in this manner. A major role of education is to "build internal controls in human beings so that they can withstand external pressures and maintain internal equilibrium" (157). We should remember that it is normal for an animal to urinate when the bladder is full and to mount any available female during the mating season, but that these behaviors may be controlled in man through training. The distinctly human quality of cerebralization of behavior is possible through education.
Human aggression may be considered a behavioral response characterized by the exercise of force with the intent to inflict damage on persons or objects. The phenomenon may be analyzed in three components: inputs, determined by environmental circumstances perceived through sensory receptors and acting upon the individual; throughputs, which are the personal processing of these circumstances through the intracerebral mechanisms established by genetic endowment and previous experiences; and outputs, represented by the expressions of individual and social behavior which constitute the observable manifestations of aggression. Increasing awareness of the need to investigate these subjects has already resulted in the creation of specialized institutes, but surprisingly enough the most essential element in the whole process of violence is usually neglected. Attention is directed to economic, ideological, social, and political factors and to their consequences, which are expressed as individual and mass behavior, while the essential link in the central nervous system is often forgotten. It is, however.
an incontrovertible fact that the environment is only the provider of sensory inputs which must be interpreted by the brain, and that any kind of behavior is the result of intracerebral activity.
It would be naive to investigate the reasons for a riot by recording the intracerebral electrical activity of the participants, but it would be equally wrong to ignore the fact that each participant has a brain and that determined neuronal groups are reacting to sensory inputs and are subsequently producing the behavioral expression of violence. Both neurophysiological and environmental factors must be evaluated, and today methodology is available for their combined study. Humanity behaves in general no more intelligently than animals would under the same circumstances, and this alarming reality is due largely to that spiritual pride which prevents men from regarding themselves and their behavior as parts of nature and as subject to its universal laws" (148). Experimental investigation of the cerebral structures responsible for aggressive behavior is an essential counterpart of social studies, and this should be recognized by sociologists as well as biologists.
In animals, the first demonstration that offensive activity could be evoked by ESB was provided by Hess (I 05), and it has subsequently been confirmed by numerous investigators. Cats under electrical stimulation of the periventricular gray matter acted "as if threatened by a dog," responding with unsheathed claws and well-aimed blows. "The animal spits, snorts or growls. At the same time the hair on its back stands on end, and its tail becomes bushy. Its pupils widen sometimes to their maximum, and its ears lie back or move back and forth to frighten the non-existing enemy" (106). In these experiments it is important to know how the cat really feels. Is it aware of its own responses? Is the hostility purposefully oriented to do harm? Or is the entire phenomenon a pseudoaffective reaction, a false or sham rage containing the motor components of offensive display without actual emotional participation? These issues have been debated over the years, but today it is clear that both sham and true rage



At upper left, the control, two friendly cats. At lower left, electrical stimulation of the anterior hypothalamus evoked an aggressive expression not directed against the other cat which, however, reacts with a defensive attitude. Above, the normal cat attacks the stimulated animal which lowers its head, flattens its ears, and does not retaliate. This experiment is an example of false rage (53).
can be elicited by ESB depending on the location of stimulation. Excitation of the anterior hypothalamus may induce a threatening display with hissing and growling which should be interpreted as false rage because, as shown in Figure 14, the display was not directed against other animals. When other cats reacted by hissing and attacking the stimulated animal, it did not retaliate or escape and simply lowered its head and flattened its ears, and these brain stimulations could not be conditioned to sensory cues.
In contrast, true rage has been demonstrated in other experiments. As shown in Figure 15, stimulation of the lateral hypothalamus produced an aggressive display clearly directed toward

Figure 15
Electrical stimulation of the lateral hypothalamus evoked true rage which is characterized by aggressive display oriented toward another cat (above); attack with well-oriented claws directed against other cats (below);


attack against investigators with whom relations had previously been friendly (above); learning of instrumental responses, such a rotating a paddle wheel, in order to stop the brain stimulation (below). In this way the cat expresses its dislike of being stimulated in a particular area (53).



a control animal which reacted properly in facing the threat. The stimulated animal started prowling around looking for fights with other subordinate animals, but avoided the most powerful cat in the group. It was evident that brain stimulation had created a state of increased aggressiveness, but it was also clear that the cat directed its hostility intelligently, choosing the enemy and the moment of attack, changing tactics, and adapting its movements to the motor reaction of its opponents Brain stimulation determined the affective state of hostility, but behavioral performance depended on the individual characteristics of the stimulated animal, including learned skills and previous experiences. Stimulations were usually tested for 5 to 10 seconds, but since it was important to know the fatigability of the effect, a longer experiment was performed, reducing the applied intensity to a level which did not evoke overt rage. The experimental subject was an affectionate cat which usually sought petting and ported while it was held in the experimenter's arms. Then it was introduced into the colony with five other cats and was radi4p stimulated continuously for two hours. During this period the animal sat motionless in a corner of the cage, uttering barely audible growls from time to time. If any other cat approached, the stimulated animal started hissing and threatening, and if the experimenter tried to pet it, the growls increased in intensity and the animal often spat and hissed. This hostile attitude disappeared as soon as the stimulation was over, and the cat became as friendly as before. These experiments demonstrated that brain excitation could modify reactions toward normal sensory stimuli and could modulate the quality of the responses in a way similar to modulation during spontaneous emotional states.
usually express their submissiveness by grimacing, crouching, and offering sexual play. In several colonies we have observed that radio stimulation of specific points in the thalamus or central gray in the boss monkey increased his aggressiveness and induced well-directed attacks against other members of the group, whom he chased around and occasionally bit, as shown in Figure 16 (56). It was evident that his hostility was oriented purposefully and according to his previous experience because he usually attacked the other male who represented a challenge to his authority, and he always spared the little female who was his favorite partner.
A high-ranking monkey expresses rage by attacking submissive members of the colony, but what would he the consequences of stimulating the brain of lower-ranking animals? Could they be induced to challenge the authority of other monkeys, including perhaps even the boss, or would their social inhibitions block the electrically induced hostility? These questions were investigated in one colony by changing its composition to increase progressively the social rank of one member, a female named Lina, who in the first grouping of four animals ranked lowest, progressing to third rank in the second group and to second rank in the third group. Social dominance was evaluated during extended control periods using the criteria of number of spontaneous agonistic and sexual interactions, priority in food getting, and territoriality. On two successive mornings in each colony Lina was radio stimulated for 5 seconds once a minute for one hour in the nucleus posterolateralis of the thalamus. In all three colonies, these stimulations induced Lina to run across the cage, climb to the ceiling, lick, vocalize, and according to her social status, to attack other animals. In group I, where l,ina was submissive, she tried to attack another monkey only once, and she was threatened or attacked 24 times. In group 2 she became more aggressive (24 occurrences) and was attacked only 3 times, while in group 3 Lina attacked other monkeys 79 times and was not threatened at all. No changes in the number of agonistic acts were observed in any group before or after the stimulation

Examples of threatening attitude and aggressive behavior produced by brain stimulation. Observe that the stimulated monkey chooses another


one as a specific target, and this animal usually expresses submissiveness by grimacing, crouching, or escaping. A toy tiger is also a suitable target for aggressive display.

hour, showing that alterations in Lina's aggressive behavior were determined by ESB.
In summary, intraspecies aggression has been evoked in cats and monkeys by electrical stimulation of several cerebral structures, and its expression is dependent on the social setting. Unlike purely motor effects including complex sequences which have no social significance, an artificially evoked aggressive act may be directed against a specific group member or may be entirely suppressed, according to the stimulated subject's social rank.
Many questions remain to be answered. Which cerebral areas are responsible for spontaneous aggressive behavior? By what mechanisms are environmental inputs interpreted as undesirable? How does cultural training influence the reactivity of specific cerebral areas? Can neurophysiological mechanisms of violence be re-educated, or are individual responses set for life after early imprinting? It is interesting that application of ESB modified the interpretation of the environment, changing the peaceful relations of a group of animals into sudden overt hostility. The same sensory inputs provided by the presence of other animals, which were neutral during control periods, were under ESB the cue for a ferocious and well-directed attack. Apparently brain stimulation introduced an emotional bias which altered interpretation of the surroundings.
While neurophysiological activity may be influenced or perhaps even set by genetic factors and past experience, the brain is the direct interpreter of environmental inputs and the determinant of behavioral responses. To understand the causes and plan remedies for intraspecific aggression in animals and man require knowledge of both sociology and neurophysiology. Electricity cannot determine the target for hostility or direct the sequences of aggressive behavior, which are both related to the past history of the stimulated subject and to his immediate adaptation to changing circumstances. Artificially triggered and spontaneously provoked aggression have many elements in common, suggesting that in both cases similar areas of the brain have been activated.
While individual and collective acts of violence may seem rather distant from the electrical discharges of neurons, we should remember that personality is not in the environment but in the nervous tissue. Possible solutions to undesirable aggression obviously will not be found in the use of ESB. This is only a methodology for investigation of the problem and acquisition of necessary information about the brain mechanisms involved. It is well known that medical treatment of cardiac patients is based on anatomical and physiological studies of the heart, and that without this information it would not have been possible to discover new drugs or to give effective medical advice. Similarly, without knowledge of the brain it will be difficult to correlate social causality with individual reactivity.
Anxiety, Fear, and Violence Evoked by ESB in Man
Anxiety has been considered the alpha and omega of psychiatry. It is one of the central themes of existential philosophy, and it shades the normal - and abnormal - life of most human beings. Several emotional states may be classified under the heading of anxiety, including fear, fright, panic, and terror, which are variations of the same basic experience. One of the most complex mental disturbances, unreasonable or excessive anxiety, including phobias and compulsive obsessions, often does not respond to standard therapies, and in some instances it has been improved by electrocoagulation of discrete areas of the frontal pole. Grey Walter (234) has claimed an 85 per cent total social recovery in a group of sixty patients with anxiety and obsessions treated with carefully dosified coagulations made through electrodes implanted in the frontal lobes.
Without entering into semantic discussions, we may consider anxiety an emotional state of conscious or subconscious tension related to real or imaginary threats to psychological or physical individual integrity. A mild degree of anxiety may mobilize, while excessive degrees may paralyze somatic and mental activity. Beyond a certain limit, anxiety has unpleasant characteristics. In normal circumstances, it is produced, as is any other emotion,
by sensory inputs from the environment and by recollections, both of which require mental elaboration of messages which may be influenced by humoral and neuronal factors. In addition, there is abundant evidence that anxiety and fear may be induced as either a primary or a secondary category of response by direct electrical stimulation of the brain. The perception or expectancy of pain can be frightening, and in some cases when ESB produced localized or generalized discomfort, patients have expressed concern about continuation of the exploratory procedures. In addition to the natural fear of possible further discomfort, there may have been a component of primary anxiety which would be difficult to evaluate.
Destruction of discrete parts of the thalamus produces relief from anxiety neurosis and obsessive-compulsive neurosis which is probably related to the interruption of tonic pathways to the frontal lobes. Stimulation of the thalamic nucleus, however, very seldom produces anxiety, and the reports of patients are limited to feelings of weakness, being different, dizziness, floating, and something like alcoholic intoxication (214).
Clearer demonstrations of direct induction of fear without any other accompanying sensations have been reported by several investigators. Lesions in the medial thalamus give effective pain relief with a minimal amount of sensory loss, and for this reason this area has often been explored electrically in cancer patients. In some cases it has produced acute anxiety attacks, which one patient vividly described as: "It's rather like the feeling of having just been missed by a car and leaped back to the curb and went B-r-r-r." Something in his guts felt very unpleasant, very unusual, and he certainly did not want to feel like that again (73). The surprising fact is that the unpleasant sensation of fear was felt in one side of the body, contralateral to the brain stimulation, Sweet (221) has reported the case of a very intelligent patient, the dean of a graduate school, who after a unilateral sympathectomy to treat his upper limb hyperhydrosis, found that his previous and customary sensation of shivering while listening to a stirring passage of music occurred in only one side and he could
not be thrilled in the sympathectomized half of his body. These cases were interesting because emotions are usually experienced in a rather diffuse and bilateral fashion unless innervation has been specifically interrupted.
The role of the thalamus in the integration of fear is also suggested by the study of a female patient whose spontaneous crippling attacks of anxiety of overwhelming intensity had led to several suicide attempts and a chronic state of depression and agitation quite refractory to drugs and psychotherapy. Stimulation of the dorsolateral nucleus of the thalamus evoked precisely the same type of attack at a level of symptomatology directly proportional to the applied intensity. It was possible to find the electrical threshold for a mild anxiety or to increase it to higher levels simply by turning the dial of the stimulator. "One could sit with one's hand on the knob and control the level of her anxiety" (73).
in one of our female patients, stimulation of a similar area in the thalamus induced a typical fearful expression and she turned to either side, visually exploring the room behind her. When asked what she was doing, she replied that she felt a threat and thought that something horrible was going to happen. This fearful sensation was perceived as real, and she had a premonition of imminent disaster of unknown cause. The effect was reliable on different days and was not altered by the use of lights and a movie camera to document the finding. Her motor activity and choice of words varied according to the environmental setting, but her facial expression and acute sensation of nonspecific, unexplainable, but real fear were similar following different stimulations. The response started with a delay of less than one second, lasted for as long as the stimulation, and did not leave observable aftereffects. The patient remembered her fear but was not upset by the memory.
Some patients have displayed anxiety and restlessness when the pallidum was stimulated at frequencies above 8 cycles per second, and they also perceived a constriction or warmth in the chest (123). A few reported a "vital anxiety in the left chest,"
and screamed anxiously if the stimulation was repeated. Intense emotional reactions have been evoked by stimulation of the amygdaloid nucleus, but responses varied in the same patient even with the same parameters of stimulation. The effect was sometimes rage, sometimes fear. One patient explained, "I don't know what came over me. I felt like air animal" (100).
The sensation of fear without any concomitant pain has also been observed as a result of ESB of the temporal lobe (230). This effect may be classified as "illusion of fear" (174) because there was obviously no real reason to be afraid apart from the artificial electrical activation of some cerebral structures. In every case, however, fear is a cerebral interpretation of reality which depends on a variety of cultural and experiential factors with logical or illogical reasons. The fact that it can be aroused by stimulation of a few areas of the brain allows the exploration of the neuronal mechanisms of anxiety, and as a working hypothesis we may suppose that the emotional qualities of fear depend on the activation of determined structures located probably in the thalamus, amygdala, and a few other as yet unidentified nuclei. This activation usually depends on the symbolic evaluation of coded sensory inputs, but the threshold for this activation may be modified-and also reached-by direct application of ESB. Knowledge of intracerebral mechanisms of anxiety and fear will permit the establishment of a more rational pharmacological and psychiatric treatment of many suffering patients, and may also help its to understand and ameliorate the increasing level of anxiety in our civilization.
It is also known that in some tragic cases, abnormal neurological processes may be the causal factor for unreasonable and uncontrollable violence. Those afflicted may often hurt or even kill either strangers or close family members usually treated with affection. A typical example was J. P., a charming and attractive 20-year-old girl with a history of encephalitis at the age of eighteen months and many crises of temporal lobe seizures and grand mal attacks for the last ten years (6o). Her main social problem was the frequent and unpredictable occurrence of rage
which on more than a dozen occasions resulted in an assault on another person such as inserting a knife into a stranger's myocardium,or a pair of scissors into the pleural cavity of a nurse. The patient was committed to a ward for the criminally insane, and electrodes Were implanted in her amygdala and hippocampus for exploration of possible neurological abnormalities. As she was rather impulsive, confinement in the EEG recording room was impractical, and she became one of the first clinical cases instrumented with a stimoceiver, which made it possible to study intracerebral activity without restraint (see Figure 4). Depth recordings taken while the patient moved freely around the ward demonstrated marked electrical abnormalities in both amygdala and hippocampus. Spontaneous periods of aimless walking coincided with an increase in the number of high-voltage sharp waves. At other times, the patient's speech was spontaneously inhibited for several minutes during which she could not answer any questions although she retained partial comprehension and awareness. These periods coincided with bursts of spike activity localized to the optic radiation (Figure 17). Transitory emotional excitement was related with an increase in the number and duration of 16-cycles-per-second bursts; but the patient read papers, conversed with other people, and walked around without causing any noticeable alterations in the telemetered intracerebral electrical activity.
During depth explorations, it was demonstrated that crises of assaultive behavior similar to the patient's spontaneous bursts of anger could be elicited by radio stimulation of contact 3 in the right amygdala. A 1.2 milliampere excitation of this point was applied while she was playing the guitar and singing with enthusiasm and skill. At the seventh second of stimulation, she threw away the guitar and in a fit of rage launched an attack against the wall and then paced around the floor for several, minutes, after which she gradually quieted down and resumed her usual cheerful behavior. This effect was repeated on two different days. The fact that only the contact located in the amygdala induced rage suggested that the neuronal field around
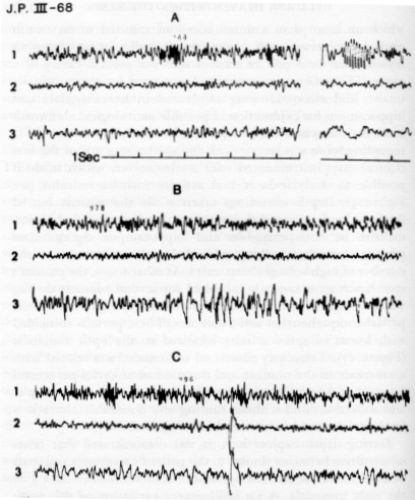
Telemetric recording of electrical activity of the brain in one of the patients shown in Figure 4. The location of the contacts was as follows: Channel i: amygdaloid nucleus; Channel 2: anterior optic radiation; Channel 3: posterior optic radiation. A: spontaneous bursts appearing in Channel i were more prominent when the patient was psychologically excited. B: sudden spontaneous arrest of speech coincided with bursts of spikes in Channel 3. C: control recordings were unmodified by friendly behavior or by different types of motor activity such as walking and reading (60).
contact 3 was involved in the patient's behavior problem, and this finding was of great clinical significance in the orientation of subsequent treatment by local coagulation.
The demonstration that amygdaloid stimulation may induce violent behavior has also been provided by other investigators. King (128) has described the case of a woman with feelings of depression and alienation, with an extremely flat tone of voice and a facial expression which was blank and unchanging during interviews, who upon stimulation of the amygdala with 5 milliamperes had greatly altered vocal inflections and an angry expression. During this time she said, "I feel like I want to get up from this chair! Please don't let me do it! Don't do this to me, I don't want to be mean!" When the interviewer asked if she would like to hit him, the patient answered, "Yeah, I want to hit something. I want to act something and just tear it up. Take it so I won't! " She then handed her scarf to the interviewer who gave her a stack of paper, and without any other verbal exchange, she tore it into shreds saying, "I don't like to feel like this." When the level of stimulation was reduced to 4 milliamperes, her attitude changed to a broad smile, and she explained, "I know it's silly, what I'm doing. I wanted to get up from this chair and run. I wanted to hit something, tear up something-anything. Not you, just anything. I just wanted to get up and tear. I had no control of myself." An increase in intensity up to 5 milliamperes again resulted in similar aggressive manifestations, and she raised her arm as if to strike.
It is notable that although the patients seemed to be out of control in these two instances of electrically induced aggression, they did not attack the interviewer, indicating that they were aware of their social situation. This finding is reminiscent of the behavior of stimulated monkeys who directed their aggressiveness according to previous experience and social rank and did not dare to challenge the authority of well-established bosses, Apparently ESB can induce a state of increased violent reactivity which is expressed in accordance with individual structure and environmental circumstances. We may conclude therefore that
artificially evoked emotional change is only one more factor in the constellation of behavioral determinants.
Pleasurable Excitation of the Animal Brain
It is surprising that in science as well as in literature more attention has been paid to suffering than to happiness. The central theme of most novels is tragedy, while happy books are hard to find; excellent monographs have been published about pain, but similar studies of pleasure are nonexistent. Typically, in the monumental Handbook of the American Physiological Society (75), a full chapter is devoted to pain, and pleasure is not even listed in the general subject index. Evidently the pursuit of happiness has not aroused as much -scientific interest as the fear of pain.
In Psychological literature the study of reward is well represented, but even there it has been considered a second-rate sensation and perhaps an artifact of a diminution of pain. It has been postulated that a truly "pleasant" sensation could not exist because organisms have a continuous tendency to minimize incoming stimuli. Pleasure was thus considered a subjective name for the diminution of drive, the withdrawal of a strong stimulation, or the reduction of pain. This "pain reduction" theory (154) has been fruitful as a basis for psychological investigations, but it is gloomy to think that we live in a 'world of punishment in which the only reality is suffering and that or brain can perceive different degrees of pain but no real pleasure. Interest in the earlier ideas of hedonism has been renewed by recent experimental studies. According to this theory, pain and pleasure are relatively independent sensations and can be evoked by different types of stimuli which are recognized by separate cerebral mechanisms. Behavior is considered to be motivated by stimuli which the organism, tries to minimize (pain) or by stimuli which the organism tries to maximize (pleasure). The brain is thought to have different systems for
the reception of these two kinds of inputs, and the psychological of pleasure or reward can be determined not only by tile state termination of pain but also by the onset of primary pleasure. The discovery of two anatomically distinct mechanisms in the brain, one for punishment, as mentioned earlier, and one for reward, provides a physiological basis for the dualistic motivation postulated in hedonism (62, 165).
The surprising fact is that animals of different species, including rats, cars, and monkeys, have voluntarily chosen to press a lever which provides electrical stimulation of specific cerebral areas. The demonstrations are highly convincing because animals which initially pressed a lever to obtain the reward of sugar pellets later pressed at similar or higher rates when electrical stimulation was substituted for food. These experiments showed conclusively that the animals enjoyed the electrical impulses which were delivered only at their own demand. Watching a rat or monkey stimulate its own brain is a fascinating spectacle. Usually each lever pressing triggers a brief 0.5-to-1.0 second brain stimulation which can be more rewarding than food. In a choice situation, hungry rats ran faster to reach tile self-stimulation lever than to obtain pellets, and they persistently pressed this lever, ignoring food within easy reach. Rats have removed obstacles, run mazes, and even crossed electrified floors to reach the lever that provided cerebral stimulation.
Not all areas of the brain involved in pleasurable effects appear equally responsive. The highest lever-pressing rates (of up to a remarkable 5,000 times per hour) were recorded by animals self-stimulating in the posterior hypothalamus; excitation of rhinencephalic structures (of only about 200 times per hour) was considered moderately rewarding; and in sensory or motor areas, animals self-stimulated at merely a chance level (of 10 to 25 times per hour), and these areas were classified as neutral. As should be expected, when stimulation was shifted from rewarding areas to nuclei in the punishment system in the same animals, they pressed the lever once and never went back,
showing that in the brain of the same animal there were two different groups of structures, one rewarding and the other aversive.
A systematic analysis of the neuroanatomical distribution of pleasurable areas in the rat (164) shows that 6o per cent of the brain is neutral, 35 per cent is rewarding, and only 5 per cent may elicit punishing effects. The idea that far more brain is involved in pleasure than in suffering is rather optimistic and gives hope that this predominance of the potential for pleasurable sensations can be developed into a more effective behavioral reality.
Because of the lack of verbal communication with animals, any ideas about what kind of pleasure, if any, may be experienced during ESB is a matter of speculation. There are some indications, however, that the perceived sensations could be related to anatomical differentiation of primary rewards of food and sex, because hungry animals self-stimulated at a higher rate in the middle hypothalamus, while administration of sexual hormones to castrated rats increased their lever pressing of more lateral hypothalamic points.
The controversial issue of how these findings in animals may relate to human behavior and the possible existence of areas involved in pleasure in the human brain has been resolved by the information obtained in patients with implanted electrodes.
Human Pleasure Evoked by ESB
On the basis of many studies during cerebral surgery, Penfield (174) has said of anger, joy, pleasure, and sexual excitement in the human brain that "so far as our experience goes, neither localized epileptic discharge nor electrical stimulation is capable of awakening any such emotion. One is tempted to believe that there are no specific cortical mechanisms associated with these emotions." This statement still holds true for the cerebral cortex, but studies in human subjects with implanted electrodes have demonstrated that electrical stimulation of the depth of the
brain can induce pleasurable manifestations, as evidenced by the spontaneous verbal reports of patients, their facial expression and general behavior, and their desire to repeat the experience. In a group of twenty-three patients suffering from schizophrenia (98), electrical stimulation of the septal region, located deep in the frontal lobes, produced an enhancement of alertness sometimes accompanied by an increase in verbal output, euphoria, or pleasure. In a more systematic study in another group of patients, further evidence was presented the rewarding effects of septal stimulation (20, 99). One man suffering from narcolepsia was provided with a small stimulator and a built-in counter which recorded the number of times that he voluntarily stimulated each of several selected points in his brain during a period of seventeen weeks. The highest score was recorded front one point in the septal region, and the patient declared that pushing this particular button made him feel "good" as if he were building up to a sexual orgasm, although he was not able to reach the end point and often felt impatient and anxious. His narcolepsia was greatly relieved by pressing this ' septal button." Another patient with psychomotor epilepsy also enjoyed septal self-stimulation, which again had the highest rate of buttton pressing and often induced sexual thoughts, Activation of the septal region by direct injection of acetylcholine produced local electrical changes in two epileptic patients and a shift in iiiood from disphoria to contentment and euphoria, usually with concomitant sexual motivation and some "orgastic sensations."
Further information was provided by another group of sixty-five patients suffering from schizophrenia or Parkinson's disease, in whom a total of 643 contacts were implanted, mainly in the anterior part of the brain (201). Results of ESB were grouped as follows: 360 points were "Positive I," and with stimulation "the patients became relaxed, at ease, had a feeling of well-being, and/or were a little sleepy." Another 31 points were "Positive II," and "the patients were definitely changed . . . in a good mood, felt good. They were relaxed, at ease, and enjoyed themselves, frequently smiling. There was a slight euphoria, but the
behavior was adequate." They sometimes wanted more stimulations. Excitation of another eight points evoked behavior classified as "Positive III," when "the euphoria was definitely beyond normal limits. The patients laughed out loud, enjoyed themselves, and positively liked the stimulation, and wanted more." ESB of another 38 points gave ambivalent results, and the patients expressed occasional pleasure or displeasure following excitation of the same area. From three other points, responses were termed "orgasm" because the patients initially expressed enjoyment and then suddenly were completely satisfied and did not want any more stimulation for a variable period of time. Finally, from about two hundred other points, ESB produced unpleasant reactions including anxiety, sadness, depression, fear, and emotional outbursts. One of the moving pictures taken in this study was very demonstrative, showing a patient with a sad expression and slightly depressed mood who smiled when a brief stimulation was applied to the rostral part of the brain, returning quickly to his usual depressed state, to smile again as soon as stimulation was reapplied. Then a ten-second stimulation completely changed his behavior and facial expression into a lasting pleasant and happy mood. Some mental patients have been provided with portable stimulators which they have used in self-treatment of depressive states with apparent clinical success.
These results indicate the need for careful functional exploration during brain surgery in order to avoid excessive euphoria or depression when positive or negative reinforcing areas are damaged. Emotional instability, in which the subject bursts suddenly into tears or laughter without any apparent reason, has been observed following some neurosurgical interventions. These major behavior problems might have been avoided by sparing the region involved in emotional regulation.
In our own experience, pleasurable sensations were observed in three patients with psychomotor epilepsy (50, 58, 109). The first case was V.P., a 36-year-old female with a long history of epileptic attacks which could not be controlled by medication.
Electrodes were implanted in her right temporal lobe and upon stimulation of a contact located in the superior part about thirty millimeters below the surface, the patient reported a pleasant tingling sensation in the left. side of her body "from my face down to the bottom of my legs." She started giggling and making funny comments, stating that she enjoyed the sensation "very much." Repetition of these stimulations made the patient more communicative and flirtatious, and she ended by openly expressing her desire to marry the therapist. Stimulation of other cerebral points failed to modify her mood and indicated the specificity of the evoked effect. During control interviews before and after ESB, her behavior was quite proper, without familiarity or excessive friendliness.
The second patient was J.M., an attractive, cooperative, and intelligent 30-year-old female who had suffered for eleven years from psychomotor and grand mal attacks which resisted medical therapy. Electrodes were implanted in her right temporal lobe, and stimulation of one of the points in the amygdala induced a pleasant sensation of relaxation and considerably increased her verbal output, which took on a more intimate character. This patient openly expressed her fondness for the therapist (who was new to her), kissed his hands, and talked about her immense gratitude for what was being done for her. A similar increase in verbal and emotional expression was repeated when the same point was stimulated on a different day, but it did not appear when other areas of the brain were explored. During control situations the patient was rather reserved and poised.
The third case was A.F., an 1 1-year-old boy with severe psychomotor epilepsy. Six days after electrode implantation in both temporal lobes, his fourth tape-recorded interview was carried out while electrical activity of the brain was continuously recorded and 5-second stimulations were applied in a prearranged sequence at intervals of about four minutes. The interviewer maintained an air of friendly interest throughout, usually without initiating conversation. After six other excitations, point LP located on the surface of the left temporal lobe was stim-
ulated for the first time, and there was an open and precipitous declaration of pleasure. The patient had been silent for the previous five-minute interval, but immediately after this stimulation lie exclaimed, "Hey! You can keep me here longer when you give me these; I like those." He went on to insist that the ongoing brain tests made him feel good. Similar statements with an emphatic expression of "feeling good" followed eight of a total sixteen stimulations of this point during the ninety-minute interview. Several of these manifestations were accompanied by a statement of fondness for the male interviewer, and the last one was accompanied by a voluptuous stretch. None of these manifestations appeared during the control prestimulation period of twenty-six minutes or during the twenty-two minutes when other points were excited. Statistical analysis of the difference between the frequency of pleasurable expressions before and after onset of stimulations proved that results were highly significant (P < 0.00 1).
The open expressions of pleasure in this interview and the general passivity of behavior could be linked, more or less intuitively, to feminine strivings. It was therefore remarkable that in the next interview, performed in a similar manner, the patient's expressions of confusion about his own sexual identity again appeared following stimulation of point LP. He suddenly began to discuss his desire to get married, but when asked, "To whom?" he did not immediately reply. Following stimulation of another point and a one-minute, twenty-second silence, the patient said, "I was thinking-there's-I was saying this to you. How to spell 'yes'-y-e-s. I mean y-o-s. No! 'You' ain't y-e-o. It's this. Y-o-u." The topic was then completely dropped. The monitor who was listening from the next room interpreted this as a thinly veiled wish to marry the interviewer, and it ivas decided to stimulate the same site again after the prearranged schedule had been completed, During the following forty minutes, seven other points were stimulated, and the patient spoke about several topics of a completely different and unrelated content. Then LP was stimulated again, and the patient started
making references to the facial hair of the interviewer and continued by mentioning pubic hair and his having been the object of genital sex play in the past. He then expressed doubt about his sexual identity, saying, "I was thinkin' if I was a boy or a girl-which one I'd like to be." Following another excitation he remarked with evident pleasure: "You're doin' it now," and then he said, "I'd like to be a girl."
In the interpretation of these results it is necessary to consider the psychological context in which electrical stimulation occurs, because the personality configuration of the subject, including both current psychodynamic and psychogenetic aspects, may be an essential determinant of the results of stimulation. Expression of feminine strivings in our patient probably was not the exclusive effect of ESB but the expression of already present personality factors which were activated by the stimulation. The balance between drive and defense may be modified by ESB, as suggested by the fact that after one stimulation the patient said without apparent anxiety, "I'd like to be a girl," but when this idea was presented to him by the therapist in a later interview without stimulation, the patient became markedly anxious and defensive. Minute-to-minute changes in personality function, influenced by the environment and by patient-interviewer relations, may modify the nature of specific responses, and these variables, which are difficult to assess, must be kept in mind.
Friendliness and Increased Conversation Under Electrical Control
Human relations evolve between the two opposite poles of love and hate which are determined by a highly complex and little understood combination of elements including basic drives, cultural imprinting, and refined emotional and intellectual characteristics. This subject has so many semantic and conceptual problems that few investigators have dared to approach it experimentally, and in spite of its essential importance, most
textbooks of psychology evade its discussion. To define friendliness is difficult although its identification in typical cases is easy, and in our daily life we are continuously evaluating and classifying personal contacts as friendly or hostile. A smiling face, attentive eyes, a receptive hand, related body posture, intellectual interest, ideological agreement, kind words, sympathetic comments, and expressions of personal acceptance are among the common indicators of cordial interpersonal relations. The expression of friendship is a part of social behavior which obviously requires contact between two or more individuals. A mutually pleasurable relation creates a history and provides each individual with a variety of optic, acoustic, tactile, and other stimuli which are received and interpreted with a "friendly bias." The main characteristic of love and friendship is precisely that stimuli coming from a favored person are interpreted as more agreeable than similar stimuli originating from other sources, and this evaluation is necessarily related to neuronal activity.
Little is known about the cerebral mechanisms of friendliness, but as is the case for any behavioral manifestation, no emotional state is possible without a functioning brain, and it may be postulated that some cerebral structures are dispensable and others indispensable both for the interpretation of sensory inputs as amicable and for the expression of friendship. Strong support for this idea derives from the fact, repeatedly proved in neurosurgery, that destruction of some parts of the brain, such as the motor and sensory cortices, produces motor deficits without modifying affective behavior, while ablation of the frontal lobes may induce considerable alteration of emotional personality. Further support has been provided by electrical stimulation of the frontal lobes, which may induce friendly manifestations.
In patient A. F., mentioned earlier in connection with pleasurable manifestations, the third inter-view was characterized by changes in the character and degree of verbal output following stimulation of one point in the temporal cortex. Fourteen
stimulations were applied, seven of them through point RP located in the inferolateral part of the right frontal lobe cortex, and the other seven through contacts located on the cortex of the right temporal lobe and depth of the left and right temporal lobes. The interview started with about five minutes of lively conversation, and during the next ten minutes the patient gradually quieted down until he spoke only about five seconds during every subsequent two-minute period. Throughout the interview the therapist encouraged spontaneous expression by reacting compassionately, by joking with, urging, and reassuring the patient, and by responding to any information offered. The attitude never produced more than a simple reply and often not even that.
In contrast to this basic situation, there were six instances of sharp increase in verbal communication and its friendly content. Each of these instances followed within forty seconds after stimulation of point RP. The only exception was the last excitation of this point when the voltage had been changed. The increases in verbal activity were rapid bat brief and without any consistency in subject material, which was typical for the patient. Qualification and quantification of the patient's conversation was made by analyzing the recorded typescript which was divided into two-minute periods and judged independently by two investigators who had no knowledge of the timing or location of stimulations. Comparison of the two-minute periods before and after these stimulations revealed a verbal increase from seventeen to eighty-eight words -and a greater number of friendly remarks, from six to fifty-three. These results were highly significant and their specificity was clear because no changes in verbalization were produced by stimulation of any of the other cerebral points. It was also evident that the evoked changes were not related to the interviewer's rather constant verbal activity. It was therefore concluded that the impressive increase in verbal expression and friendly remarks was the result of electrical stimulation of a specific point on the cortex of the temporal lobe.